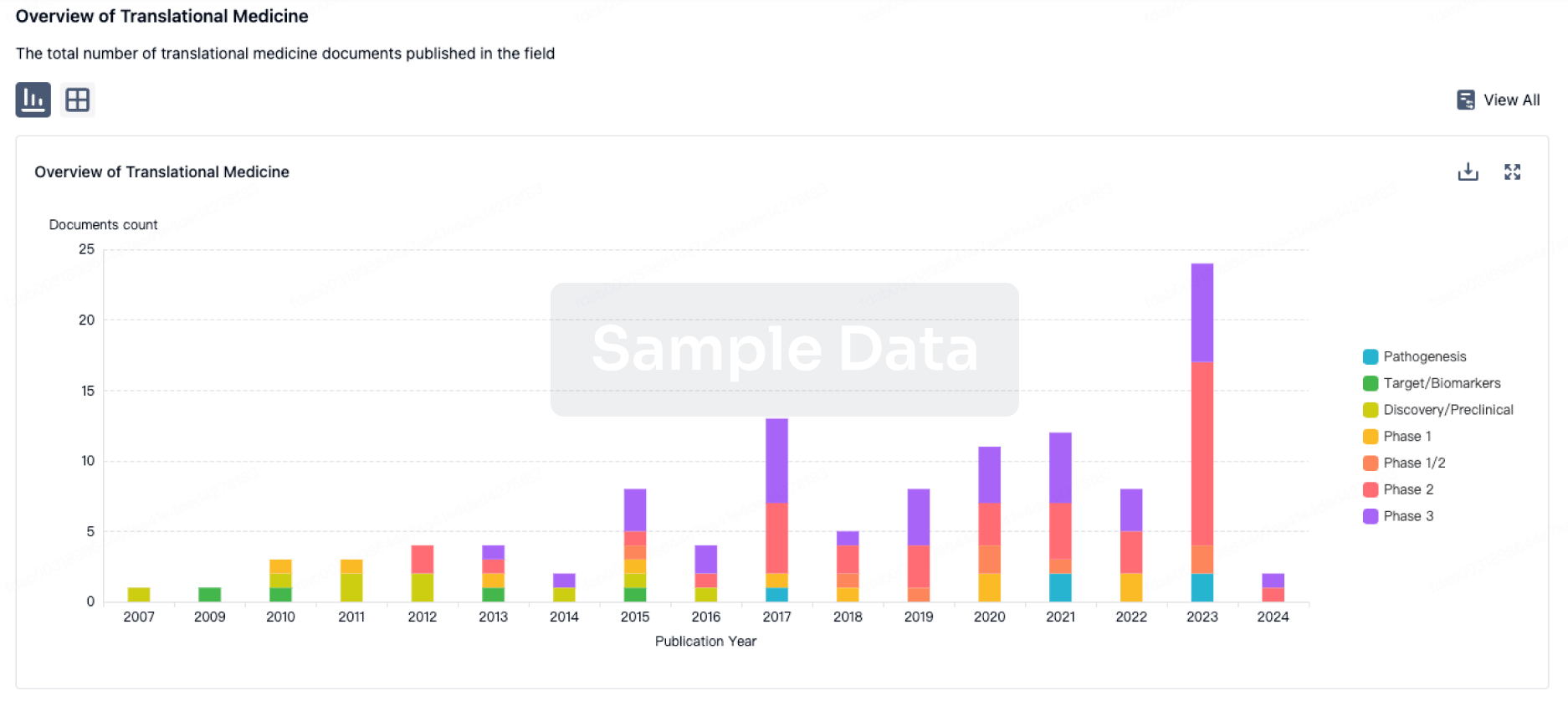
Human brown adipose tissue (BAT) is experimentally modeled to better understand the biology of this important metabolic tissue, and also to enable the potential discovery and development of novel therapeutics for obesity and sequelae resulting from the persistent positive energy balance. This chapter focuses on translation into humans of findings and hypotheses generated in nonhuman models of BAT pharmacology. Given the demonstrated challenges of sustainably reducing caloric intake in modern humans, potential solutions to obesity likely lie in increasing energy expenditure. The energy-transforming activities of a single cell in any given tissue can be conceptualized as a flow of chemical energy from energy-rich substrate molecules into energy-expending, endergonic biological work processes through oxidative degradation of organic molecules ingested as nutrients. Despite the relatively tight coupling between metabolic reactions and products, some expended energy is incidentally lost as heat, and in this manner a significant fraction of the energy originally captured from the environment nonproductively transforms into heat rather than into biological work. In human and other mammalian cells, some processes are even completely uncoupled, and therefore purely energy consuming. These molecular and cellular actions sum up at the physiological level to adaptive thermogenesis, the endogenous physiology in which energy is nonproductively released as heat through uncoupling of mitochondria in brown fat and potentially skeletal muscle. Adaptive thermogenesis in mammals occurs in three forms, mostly in skeletal muscle and brown fat: shivering thermogenesis in skeletal muscle, non-shivering thermogenesis in brown fat, and diet-induced thermogenesis in brown fat. At the cellular level, the greatest energy transformations in humans and other eukaryotes occur in the mitochondria, where creating energetic inefficiency by uncoupling the conversion of energy-rich substrate molecules into ATP usable by all three major forms of biological work occurs by two primary means. Basal uncoupling occurs as a passive, general, nonspecific leak down the proton concentration gradient across the membrane in all mitochondria in the human body, a gradient driving a key step in ATP synthesis. Inducible uncoupling, which is the active conduction of protons across gradients through processes catalyzed by proteins, occurs only in select cell types including BAT. Experiments in rodents revealed UCP1 as the primary mammalian molecule accounting for the regulated, inducible uncoupling of BAT, and responsive to both cold and pharmacological stimulation. Cold stimulation of BAT has convincingly translated into humans, and older clinical observations with nonselective 2,4-DNP validate that human BAT's participation in pharmacologically mediated, though nonselective, mitochondrial membrane decoupling can provide increased energy expenditure and corresponding body weight loss. In recent times, however, neither beta-adrenergic antagonism nor unselective sympathomimetic agonism by ephedrine and sibutramine provide convincing evidence that more BAT-selective mechanisms can impact energy balance and subsequently body weight. Although BAT activity correlates with leanness, hypothesis-driven selective β3-adrenergic agonism to activate BAT in humans has only provided robust proof of pharmacologic activation of β-adrenergic receptor signaling, limited proof of the mechanism of increased adaptive thermogenesis, and no convincing evidence that body weight loss through negative energy balance upon BAT activation can be accomplished outside of rodents. None of the five demonstrably β3 selective molecules with sufficient clinical experience to merit review provided significant weight loss in clinical trials (BRL 26830A, TAK 677, L-796568, CL 316,243, and BRL 35135). Broader conclusions regarding the human BAT therapeutic hypothesis are limited by the absence of data from most studies demonstrating specific activation of BAT thermogenesis in most studies. Additionally, more limited data sets with older or less selective β3 agonists also did not provide strong evidence of body weight effects. Encouragingly, β3-adrenergic agonists, catechins, capsinoids, and nutritional extracts, even without robust negative energy balance outcomes, all demonstrated increased total energy expenditure that in some cases could be associated with concomitant activation of BAT, though the absence of body weight loss indicates that in no cases did the magnitude of negative energy balance reach sufficient levels. Glucocorticoid receptor agonists, PPARg agonists, and thyroid hormone receptor agonists all possess defined molecular and cellular pharmacology that preclinical models predicted to be efficacious for negative energy balance and body weight loss, yet their effects on human BAT thermogenesis upon translation were inconsistent with predictions and disappointing. A few new mechanisms are nearing the stage of clinical trials and may yet provide a more quantitatively robust translation from preclinical to human experience with BAT. In conclusion, translation into humans has been demonstrated with BAT molecular pharmacology and cell biology, as well as with physiological response to cold. However, despite pharmacologically mediated, statistically significant elevation in total energy expenditure, translation into biologically meaningful negative energy balance was not achieved, as indicated by the absence of measurable loss of body weight over the duration of a clinical study.