Request Demo
Last update 08 May 2025
BRPF2 x ATAD2
Last update 08 May 2025
Related
Target |
Mechanism |
Active Org. |
Originator Org. |
Active Indication |
Inactive Indication |
Drug Highest Phase |
First Approval Ctry. / Loc. |
First Approval Date |
100 Clinical Results associated with BRPF2 x ATAD2
Login to view more data
100 Translational Medicine associated with BRPF2 x ATAD2
Login to view more data
Login to view more data
01 Mar 2018Journal of Cellular PhysiologyQ2 · BIOLOGY
Biological function and histone recognition of family IV bromodomain‐containing proteins
Q2 · BIOLOGY
Review
Author: Glass, Karen C. ; Lloyd, Jonathan T.
26 Oct 2017Chemistry – A European JournalQ2 · CHEMISTRY
Synthesis and Demonstration of the Biological Relevance of sp3‐rich Scaffolds Distantly Related to Natural Product Frameworks
Q2 · CHEMISTRY
Article
Author: Nelson, Adam ; Foley, Daniel J. ; Doveston, Richard G. ; Aimon, Anthony ; Talon, Romain ; Marsden, Stephen P. ; Collins, Patrick M. ; von Delft, Frank ; Craven, Philip G. E. ; Churcher, Ian
01 Dec 2014Epigenetics & ChromatinQ2 · BIOLOGY
Assessing cellular efficacy of bromodomain inhibitors using fluorescence recovery after photobleaching
Q2 · BIOLOGY
ArticleOA
Author: Jean-Philippe Lambert ; Martin Philpott ; Clarence Yapp ; Catherine M Rogers ; Chris Wells ; Anne-Claude Gingras ; Stefan Knapp ; Claire Strain-Damerell ; Nicola A Burgess-Brown ; Susanne Müller
Analysis
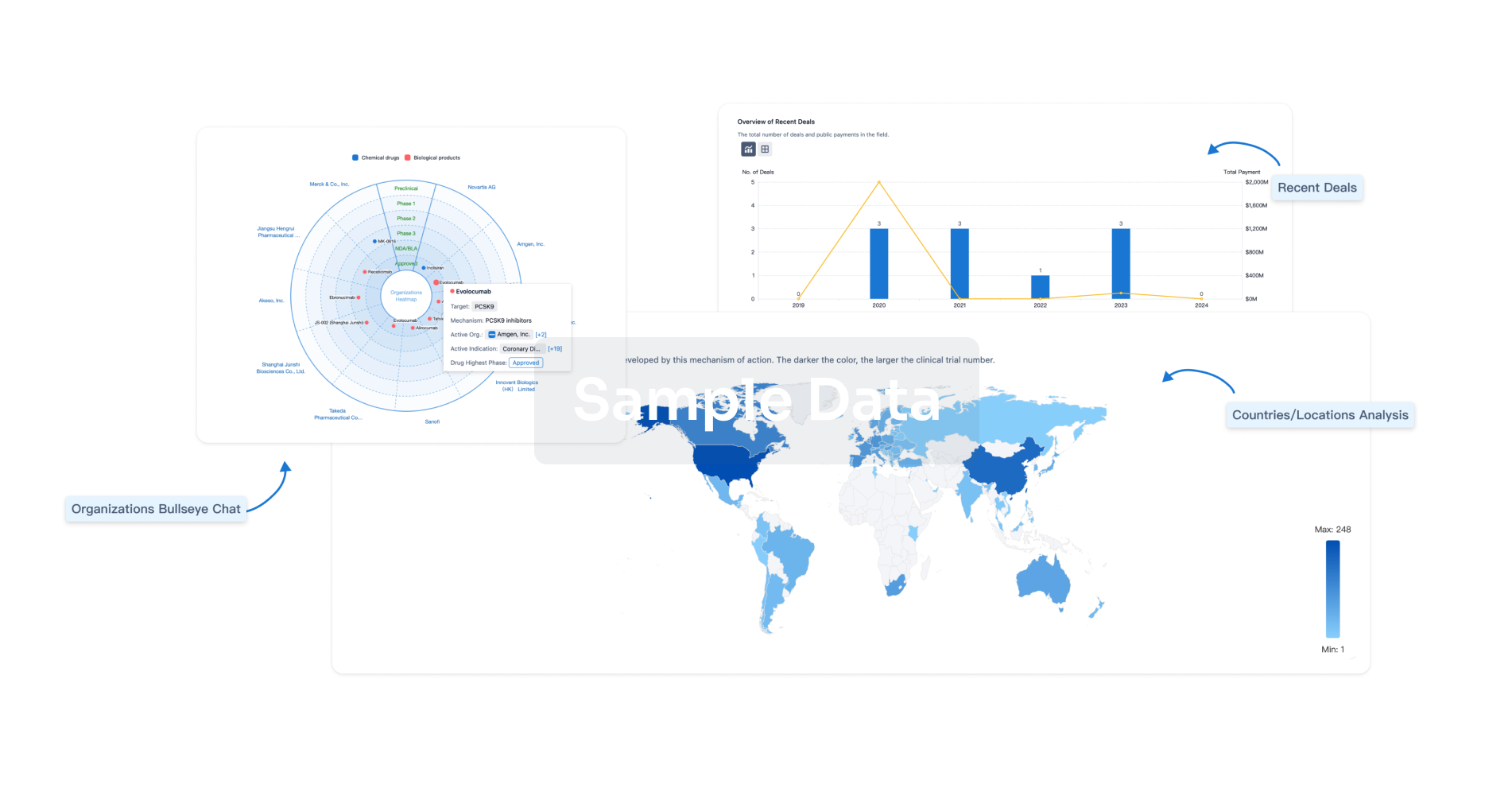
Perform a panoramic analysis of this field.
login
or
AI Agents Built for Biopharma Breakthroughs
Accelerate discovery. Empower decisions. Transform outcomes.
Get started for free today!
Accelerate Strategic R&D decision making with Synapse, PatSnap’s AI-powered Connected Innovation Intelligence Platform Built for Life Sciences Professionals.
Start your data trial now!
Synapse data is also accessible to external entities via APIs or data packages. Empower better decisions with the latest in pharmaceutical intelligence.
Bio
Bio Sequences Search & Analysis
Sign up for free
Chemical
Chemical Structures Search & Analysis
Sign up for free